Merci à Manon LEPRAT d’accepter que nous reprenions ici une partie de son travail pour vous informer.
La peau - Définitions / Problèmatiques / Maladies
La peau en quelques mots
Quelques notions scientifiques, sur la peau.
Les informations qui suivent sont extraits de la thèse de Manon LEPRAT, intitulée "Le Vitiligo: de la physiopathologie aux traitements" et présentée le 18/05/2015 à la facutlé de pharmacie de l'Université de Clermont-Ferrand 1.
Jean-Marie Meurant
Président de la Fédération Française de la Peau
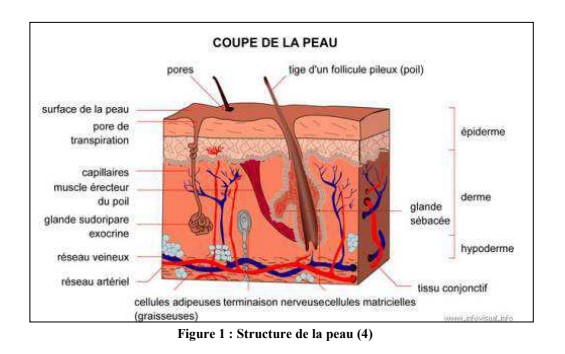
I. Structure de la peau
La peau est une enveloppe, qui recouvre notre corps. Elle représente 1/3 du poids du corps humain et possède une surface d’environ 2 m2 chez un adulte.
La protection, les mécanismes de régulation thermique et métaboliques, la cicatrisation et le système sensoriel font partie des grandes fonctions de la peau. La peau est le témoin du dysfonctionnement de certains organes.
1. Une structure stratifiée
La peau est un organe complexe, subdivisé en 3 couches superposées qui vont de la superficie vers la profondeur : l’épiderme, le derme et l’hypoderme (figure 1). L’ensemble de la peau est séparé des organes sous-jacents par le fascia.
a. L’épiderme
L’épiderme, ou couche superficielle, protège l’organisme contre les agressions extérieures. Il forme, suivant la migration des kératinocytes : un épithélium de revêtement, stratifié, pavimenteux et kératinisé.
Son épaisseur est variable selon la localisation et est déterminée par l’épaisseur de la couche cornée.
L’épiderme est en renouvellement constant, il met en moyenne 21 jours pour se renouveler. Le renouvellement repose sur un processus de différenciation précis, partant de la base de l’épiderme vers la surface.
Une protéine fibreuse est produite à ce niveau, il s’agit de la kératine, très résistante, elle a un rôle de barrière.
L’épiderme est séparé du derme par la membrane basale.
Il n’est pas vascularisé (vaisseaux sanguins ou lymphatiques), il contient cependant des terminaisons nerveuses sensitives.
Il est constitué essentiellement de kératinocytes.
L’épiderme est formé de 4 assises cellulaires (figure 2) :
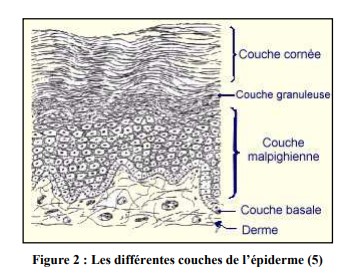
i. La couche basale ou couche germinative (ou stratum germinativum)
Elle est formée d’une assise de cellules cubiques, implantées sur les papilles du derme superficiel (directement en contact avec la jonction dermo-épidermique).
Elle est composée en grande partie de kératinocytes, entre lesquels sont insérés quelques mélanocytes.
C’est le siège des divisions cellulaires : chaque cellule basale se divise par mitose en deux kératinocytes, l’un reste au niveau de la couche basale pour se diviser et le second migre vers les couches superficielles pour ensuite se différencier.
ii. La couche épineuse ou corps muqueux de Malpighi (ou stratum spinosum)
Elle est formée d’environ cinq à six couches de cellules polygonales.
Des cellules sont hérissées de spicules ou « épines », d’où le nom de couche épineuse. Ces épines appelées desmosomes, sont des moyens de fixation intercellulaires.
Les desmosomes ont la faculté de synthétiser des protéines cytoplasmiques, notamment de la kératine.
La présence de mélanocytes à ce niveau est le signe d’un début de synthèse de mélanines.
iii. La couche granuleuse (ou stratum granulosum)
Elle est constituée d’environ trois à cinq couches de cellules granuleuses, aplaties, fusiformes et parallèles.
Le cytoplasme contient des granules de kératohyaline, qui agglutinent les fibres de kératine entre elles. Cette granulation basophile est à l’origine de l’appellation de la couche granuleuse. L’assise cellulaire est plus épaisse et rigide. Les noyaux ont tendance à régresser.
iv. La couche cornée (ou stratum corneum)
Il s’agit de la couche la plus superficielle de l’épiderme. Elle s’organise par la superposition de cellules plates, compactes, à membrane épaisse, anuclées, complètement kératinisées : définissant les cornéocytes.
Le nombre de couches de cornéocytes est variable selon les régions du corps (15 à 20 au niveau du dos et quelques centaines au niveau de la plante des pieds). Les cornéocytes sont en desquamation perpétuelle.
v. Les cellules de l’épiderme
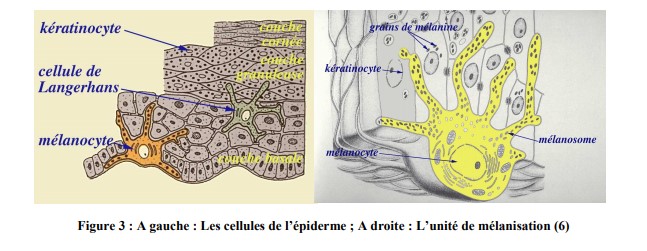
Il existe quatre principaux types de cellules épidermiques (figure 3) :
– Les kératinocytes (80%) :
D’origine ectodermique, ils sont présents dans tout l’épiderme. Trois grandes fonctions leur sont attribuées : cohésion de l’épiderme, barrière mécanique et physique. Ils contribuent au maintien de la cohésion de l’épiderme, par l’intermédiaire de leur cytosquelette et leur système de jonction pour établir des liens entre eux et la matrice extracellulaire.
Ils assurent la protection de l’épiderme contre les agressions mécaniques par l’action d’un processus de différenciation épidermique terminal.
Grâce aux mélanosomes de type IV, phagocytés à partir des mélanocytes, ils font barrière face aux radiations lumineuses.
Leur migration de la couche basale à la couche cornée s’effectue normalement en 3 à 4 semaines.
– Les mélanocytes (13%), la deuxième grande population de l’épiderme :
Ils proviennent des crêtes neuronales.
Ils sont localisés principalement dans la couche basale concernant la population blanche et au niveau de tout l’épiderme pour la population noire.
Ils sont le lieu de synthèse des mélanines, produites dans des organites spécialisés, appelés les mélanosomes. De plus, le mélanocyte se distingue par un cytoplasme clair, un appareil de Golgi et un réticulum endoplasmique mature.
La mélanine est le pigment responsable de la coloration de la peau, elle est créée à partir de la tyrosine, les grains de mélanine sont stockés dans le cytoplasme des mélanocytes.
Leur forme étoilée ainsi que la présence de prolongements s’étendant entre les
kératinocytes permettent le transfert des grains de mélanine.
– Les cellules de Langerhans (2 à 4 %) :
Elles sont situées au niveau de la couche épineuse.
Issues de la moelle hématopoïétique, elles migrent vers l’épiderme pour capturer les exoantigènes et les transformer. Ensuite, elles migrent vers le derme et les vaisseaux lymphatiques, où elles vont exprimer les antigènes avec les molécules du CMH (Complexe Majeur d’Histocompatibilité) de classe II.
Ce sont des cellules immunocompétentes, elles sont capables de capter des antigènes ayant pénétré dans l’organisme, elles interagissent avec les lymphocytes T CD4+ (Cluster de différentiation) au cours de la réponse immunitaire.
En microscopie électronique, les cellules de Langerhans apparaissent claires et sont caractérisées par la présence de granules de Birbeck, en forme de raquette de tennis.
– Les cellules de Merkel (1 à 2 %) :
Ces cellules sont présentes au niveau de la couche basale et proviennent des crêtes neuronales. Elles sont impliquées dans les fonctions inductives et trophiques des terminaisons nerveuses. Ce sont des récepteurs du sens du toucher et du tact.
En microscopie électronique, les cellules de Merkel apparaissent entre les kératinocytes basaux, à proximité d’une terminaison nerveuse. Des « vésicules à cœur dense » dans le cytoplasme leur sont caractéristiques.
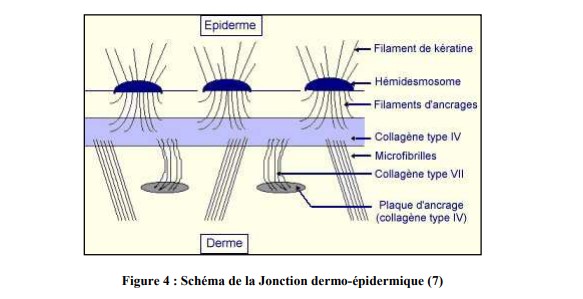
b. La jonction dermo-épidermique
La jonction dermo-épidermique, essentiellement constituée par la membrane basale, sépare comme son nom l’indique, le derme de l’épiderme. Il s’agit d’une membrane fine d’1 µm.
En microscopie optique, après des techniques de colorations spécifiques, la jonction dermoépidermique apparait comme une ligne ondulée, définie par une alternance de « crêtes épidermiques » dans le derme et de « papilles dermiques » dans l’épiderme.
En microscopie électronique, sa structure lamellaire comprend 3 régions : des cellules basales de l’épiderme, la lamina lucida claire aux électrons et la lamina densa dense aux électrons. Sa structure est beaucoup plus complexe au niveau des points d’ancrage (figure 4).
Des études d’immuno-histochimie ont mis en évidence l’existence de constituants spécialisés impliqués dans le maintien de l’intégrité de la jonction dermo-épidermique. On peut citer notamment :
- L’antigène BP (Bullous Pemphigoid) 230 : au point d’ancrage des tonofilaments des hémidesmosomes ;
- L’intégrine a6b4 et l’antigène BP 180 : molécules transmembranaires des
hémidesmosomes ; - La laminine 5 : au niveau des filaments d’ancrage ;
- Le collagène VII : au niveau des fibrilles d’ancrage.
c. Le derme
Le derme est un tissu de soutien et de nutrition pour l’épiderme. Le derme est un tissu conjonctif complexe, à la différence de l’épiderme, il est richement vascularisé. Son épaisseur est plus importante que celle du l’épiderme (0,5 à 2 mm contre 0,1 mm). Le derme confère élasticité et tonicité à la peau de part la présence de protéines fibreuses.
i. Les cellules
Ce sont principalement les fibroblastes, ils synthétisent les constituants de la matrice extracellulaire. Les adipocytes jouent un rôle dans le métabolisme cellulaire. Les histiocytes agissent au cours du phénomène de la cicatrisation. Le derme est pourvu de cellules migrantes comme les cellules du système immunitaire (mastocytes, lymphocytes…), les cellules sanguines.
ii. La matrice intercellulaire
Elle est composée de protéines fibreuses baignant dans la substance fondamentale (composée d’acide hyaluronique et de protéoglycanes). On décompte trois types de fibres :
- Les fibres de collagène : groupées en faisceaux parallèles, elles représentent 95% des protéines du derme et en forment la trame. Le collagène donne au derme sa solidité et sa résistance à la traction.
- Les fibres d’élastine : ces fibres élastiques, organisées en réseau, apportent souplesse à la peau. Ce réseau élastique se compose de fibres d’élaunine et d’oxytalanes.
- Les fibres de réticuline : elles enveloppent les faisceaux de collagène, elles servent de support au derme.
La substance fondamentale ou gel interfibrillaire, remplit les interstices entre les différentes fibres. Ce gel amorphe est constitué d’eau, de sels minéraux, de glycoprotéines de structure comme la fibronectine, de glycosaminoglycanes (macromolécules formées de mucopolysaccharides et de protéines). Le gel permet de retenir l’eau de la matrice, assurant ainsi la tonicité de la peau.
On distingue deux types de derme en fonction de la densité de ces fibres :
- Le derme papillaire ou superficiel : formé d’un tissu conjonctif lâche composé de fines fibres de collagène ;
- Le derme réticulaire (moyen et profond) : le tissu conjonctif est dense, dessiné par un réseau à maille serrée constitué de fibres de collagène denses et des fibres d’élastines.
d. L’hypoderme ou tissu sous-cutané
Il n’y a pas de limite franche entre le derme et l’hypoderme.
L’hypoderme sépare le derme des muscles et des os sous-jacents. Il est absent au niveau des oreilles, des yeux et des organes génitaux masculins.
Il est formé d’un tissu conjonctif lâche contenant des cellules adipeuses et graisseuses. Son épaisseur est très variable selon les sujets, l’âge, la localisation corporelle et la nutrition.
Le pannicule adipeux est réparti différemment chez l’homme et chez la femme. Chez l’homme, il prédomine au niveau des parties supérieures du corps, précisément au-dessus de l’ombilic vers l’abdomen. Chez la femme, il se localise principalement dans la partie inférieure du corps, au dessous de l’ombilic.
i. Structure
L’hypoderme est composé de lobes graisseux, limités par des travées de fibres de collagènes issues du derme. Les parois ou septums interlobulaires conjonctivo-élastiques séparent les lobules graisseux entre eux permettant ainsi le passage des nerfs et des vaisseaux à destination du derme. Les lobes graisseux sont subdivisés en lobules graisseux, comprenant des logettes
remplies d’adipocytes.
ii. Rôles
L’hypoderme a de multiples fonctionnalités dont principalement un rôle de protection vis-àvis des chocs mécaniques et thermiques et de réserve d’énergie et de nutriments.
- Action métabolique : les adipocytes mettent en réserve les lipides sous formes de triglycérides et fournissent des acides gras en cas de demande énergétique.
- Action plastique : l’hypoderme modèle la silhouette en fonction de l’âge, de sexe, de l’état nutritionnel.
- Protection mécanique : l’hypoderme est un « pare-choc », il possède des
récepteurs de Vater-Pacini, appelés également corpuscules de Pacini, ce sont des mécanorécepteurs sensoriels sensibles aux pressions et aux vibrations. - Thermorégulation : le tissu adipeux est isolant.
2. Les organes annexes
Le follicule pilo-sébacé se compose :
- Du poil : structure kératinique morte sécrétée par le follicule pileux. Il a une fonction thermorégulatrice.
- La glande sébacée : sécrète un sébum riche en acides gras s’émulsionnant avec la sueur pour former un film hydrolipidique assurant l’imperméabilité de l’épiderme, le protégeant ainsi de la déshydratation.
On distingue deux types de glandes sudoripares :
- Les glandes eccrines agissent dans le phénomène de thermorégulation.
- Les glandes apocrines sécrètent la sueur.
II. Physiologie de la pigmentation
Notre couleur de peau résulte d’une alliance subtile de pigments mélaniques issus de la production mélanocytaire. La pigmentation cutanée représente un élément majeur de défense de l’organisme, par sa protection contre les effets délétères des rayons ultraviolets.
La pigmentation de la peau, des poils, des yeux, varie en fonction de la qualité et de la quantité des pigments mélaniques. Des variations de couleurs peuvent également être engendrées par un épaississement de l’épiderme.
La mélanogenèse est un processus complexe sous l’influence de facteurs génétiques, chimiques mais aussi d’agents extérieurs.
1. La mélanogenèse
Elle correspond à la synthèse des mélanines, elle détermine la fonction principale de cellules spécialisées appelées les mélanocytes (figure 5).
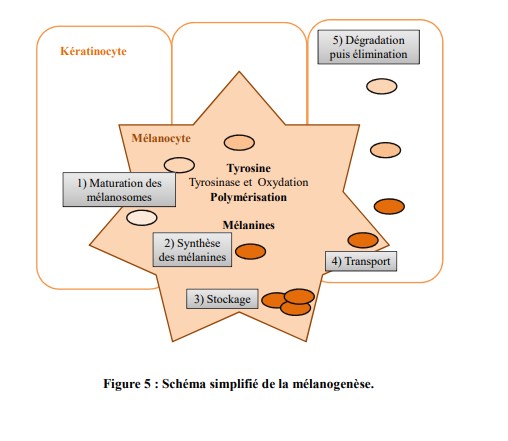
a. Les mélanines
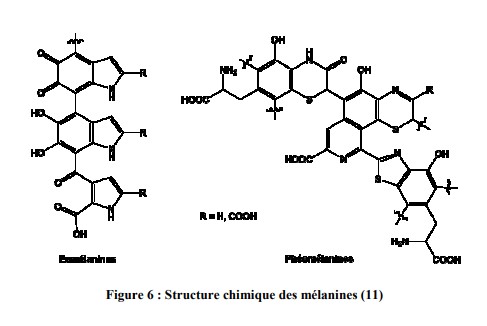
Deux familles de mélanines sont produites suivant leur structure chimique (figure 6) et leur couleur : les eumélanines et les phaeomélanines sont réparties en proportions différentes chez l’homme.
i. Les eumélanines
Les eumélanines de couleur brune ou noire, sont des mélanines de haut poids moléculaire, insolubles dans la plupart des solvants et sont pauvres en soufre. Elles forment un corps chimique absorbant totalement la lumière.
Ce corps chimique résulte de la polymérisation de nombreux radicaux phénols oxydés en quinone puis d’une cyclisation. Elles sont prédominantes chez les sujets de phototype foncé.
ii. Les phaeomélanines
Les phaeomélanines de couleur jaune-orangé, sont solubles en milieu basique. Elles sont issues de la polymérisation oxydative des cystéinyldopas, ce sont des composés soufrés et azotés. Elles sont prédominantes chez les sujets de phototype clair.
iii. Leur synthèse
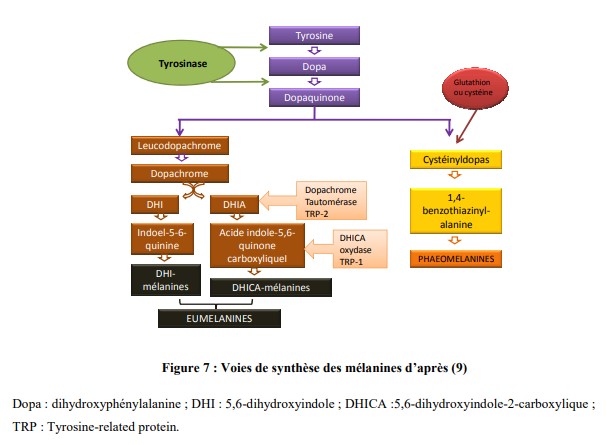
Ces mélanines proviennent d’une réaction enzymatique. La tyrosine est transformée en dihydroxyphénylalanine puis est oxydée en dopaquinone par l’intermédiaire de la tyrosinase.
Cette tyrosinase est l’enzyme limitante de cette voie de synthèse.
Ensuite, à partir de la dopaquinone, deux types de voies divergent, celle de l’eumélanogenèse et celle de la phaeomélanogenèse.
- La synthèse des eumélanines nécessite la présence de trois enzymes : la
tyrosinase-related protein-1 (TYRP1), la tyrosinase-related protein-2 (TYRP2) et la dopachrome tautomérase (Dct). - La synthèse des phaeomélanines requiert l’incorporation de dérivés soufrés tels que le glutathion ou la cystéine.
Le ratio eumélanine/phaeomélanine enzymes et la disponibilité en tyrosine ou en composés soufrés. Ce rapport définit la couleur constitutive de la peau d’une personne, il peut varier également chez un même individu, en
particulier sous l’influence des rayons été démontré récemment que la lumière visible aurait un effet les phototypes foncés.
iv. Action des rayons
Sous l’effet des rayons UV, la synthèse des mélanines s’accroit et leur transfert vers les kératinocytes est accéléré. La production des pigments mélaniques est une réponse adaptative de l’organisme face à des expositions prolongées au soleil. Cette pigmentation traduit la capacité de l’individu à développer un bronzage, mécanisme de protection naturel de la peau.
Les mélanines forment un filtre pour les rayonnements visibles et UV, elles définissent un système photoprotecteur important.
Les eumélanines sont dotées de phaeomélanines. Les eumélanines absorbent les photons émis et captent les cellules par les radiations UV, elles empêchent ainsi que l’ADN (Acide DésoxyriboNucléique) soit endommagé.
Les phaeomélanines à la différence des eumélanines, participeraient indépendamment ou non des radiations UV, à la genèse de mélanomes par des mécanismes l’ADN (elles ont donc plutôt un rôle carcinogène).
b. La mélanocytogenèse
Les mélanocytes dérivent pour la majeure partie de la crête neuronale. Les mélanocytes fonctionnels sont obtenus lors de l’embryogenèse où les cellules de la crête neuronale subissent une cascade de stimulations. Certaines molécules actrices de la mélanocytogenèse ont pu être identifiées. On peut citer :
- Le microphthalmia-associated transcription factor (MITF) : c’est en particulier le facteur de transcription des enzymes intervenant dans la synthèse des mélanines, mais il permet aussi la survie des mélanocytes en régulant le B-cell-lymphoma-2 (Bcl2).
- Le paired box gene 3 (PAX-3) et SRY-box containing gene 10 (SOX-10) sont impliqués dans la différentiation des mélanocytes.
Ces trois agents sont étroitement liés entre eux de part leur expression et leur action dans la chaîne réactionnelle.
Sur la surface des mélanocytes est localisé un récepteur à la tyrosine kinase appelé « c-KIT », son ligand le f(SCF) est synthétisé par les kératinocytes. Ces deux facteurs interviennent lors de la prolifération des mélanocytes et contribuent aussi à leur survie.
Des interactions apparaissent entre le récepteur c-KIT et MITF au cours de leur phosphorylation à l’origine des différentes voies de réactions.
D’autres facteurs agissent également dans la différentiation des mélanocytes, on peut décrire notamment : un facteur de transcription spécifique de la crête neuronale, le récepteur de l’endothéline B (EDNRB) et son ligand l’endothéline 3 (EDN3).
Les gènes donnant naissance à ces multiples facteurs peuvent subir des mutations et engendrer des troubles pigmentaires en particulier des hypomélanoses, que l’on décrira par la suite.
c. La mélanogenèse
La synthèse de mélanines est un processus complexe impliquant plusieurs enzymes pour obtenir les pigments mélaniques (figure 7).
Les principales enzymes identifiées sont au nombre de trois : la tyrosinase, la TYRP1 et la TYRP2.
La L-tyrosine, un acide aminé soufré, est capturée par le mélanocyte dans le sang circulant afin d’être par la suite incorporée dans le mélanosome.
On observe près de 40% de séquences d’acides aminés homologues au niveau de la structure de ces enzymes, notamment en ce qui concerne la région fonctionnelle. Ces enzymes n’ont cependant pas les mêmes activités catalytiques et sont codées par des gènes distincts.
Faites un don
Faire un don c'est avant tout faire avancer la recherche et aider les autres.
Devenir bénévole
Devenez bénévoles dans l'une des associations adhérentes.
Participez aux événements
Chaque association propose des événements à suivre avec attention.
Suivez nos actualités
Suivez les actualités des associations adhérentes et de la Fédération.